

Abstract
Sickle cell disease (SCD) is caused by a 20A>T mutation in the β-globin gene, and can be cured by therapeutic β-globin gene addition into hematopoietic stem cells (HSCs) with lentiviral transduction. However, this method relies upon random integration, leaving the SCD mutation intact and potentially inducing insertional mutagenesis. Genome editing technologies have the potential to correct the SCD mutation without integration, producing adult hemoglobin (Hb) while simultaneously eliminating sickle Hb. In this study, we investigated CRISPR/Cas9-based gene correction for SCD CD34+ cells.
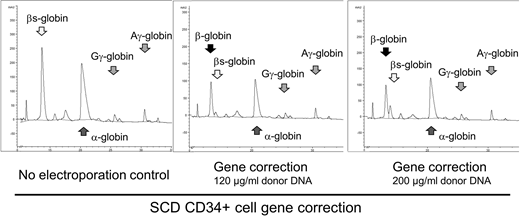
Plerixafor-mobilized SCD CD34+ cells were transfected by electroporation using the GMP-compliant, FDA Master File-supported, and scalable MaxCyte GT System to deliver SCD mutation-specific guide RNA at 200mg/ml, SpCas9 mRNA at 200mg/ml or protein at 120mg/ml, and single strand donor DNA with a normal β-globin sequence at 80, 120, or 200mg/ml. We chose Cas9 mRNA and single strand donor DNA due to the ease of clinical grade large-scale production and to avoid the need for viral vector manufacturing. Following erythroid differentiation, gene correction efficiency was evaluated at DNA levels by deep sequencing and at protein levels by reverse-phase HPLC. Cell viability was reduced to 76-87% after electroporation, compared to 90% in the control. We observed high-efficiency genome editing (29-34% gene correction and 49-58% indels) with Cas9 mRNA, showing donor DNA concentration dependence, and editing levels were comparable to Cas9 protein (39% correction and 43% indels). 15-23% Biallelic and 17-26% monoallelic gene correction were detected at the clonal level by colony assay. After erythroid differentiation, up to 54% normal β-globin production was observed with Cas9 mRNA (Figure), comparable to Cas9 protein (67%), while βs-globin amounts were markedly reduced under both conditions (6-10%). Similar correction efficiencies were obtained from two additional SCD patients' CD34+ cells at DNA levels (28-35%) and protein levels (33-56%). These data demonstrate that Cas9 mRNA and single strand donor DNA allow for efficient gene correction in SCD CD34+ cells, exceeding the therapeutic threshold of 20% in SCD.
We then evaluated off-target effects on the δ-globin gene, which was reported as a major off-target site in β-globin gene editing due to high homology; however, almost no off-target effects (0.6-1.3% indels) were detected. Interestingly, gene conversion in the 9T>C polymorphism (11bp upstream of SCD mutation) on the β-globin gene was observed, and this conversion always occurred with SCD gene correction (26-33% of SCD gene correction), suggesting that gene conversion is strongly affected by distance from the target site.
In addition, we evaluated genome editing among subpopulations of CD34+ cells from 3 healthy donors under the same conditions (normal β-globin to SCD mutation). We observed similar editing efficiencies (conversion and indels) among more immature (CD34+CD133+CD90+) and relatively differentiated populations (CD34+CD133+CD90-, CD34+CD133-, and CD34-) as well as among cells at different phases of the cell cycle (G0/G1, S, and G2/M), suggesting that similar gene correction efficiencies are obtained in all CD34+ cell populations, including the HSC population. We have begun efforts to evaluate gene-corrected SCD CD34+ cell engraftment in the mouse xenograft model, as similarly corrected X-CGD CD34+ cells were engrafted in immunodeficient mice.
To examine the effects of indels in the β-globin gene, we next evaluated Hb production from genome-edited SCD CD34+ cells (2 patients) without donor DNA. Editing without donor DNA resulted in 63-70% indels (compared to 26-29% correction and 46-53% indels with donor DNA) and increased non-adult Hb production (small amounts of fetal Hb and significant amounts of a Hb variant), which will require further investigation to characterize.
In summary, we observed efficient gene correction in SCD CD34+ cells with a simple Cas9 mRNA, single strand donor DNA, and guide RNA method, resulting in ~30% gene correction and ~50% indels. After erythroid differentiation, the majority of Hb detected was adult Hb; we detected up to 54% normal β-globin production with a marked reduction of βs-globin to ~10%. Evaluation of engraftment potential is required for gene-corrected CD34+ cells, but these methods would be clinically applicable for gene correction in SCD.
Li:MaxCyte, Inc.: Employment. Allen:MaxCyte, Inc.: Employment. Peshwa:MaxCyte, Inc.: Employment.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal